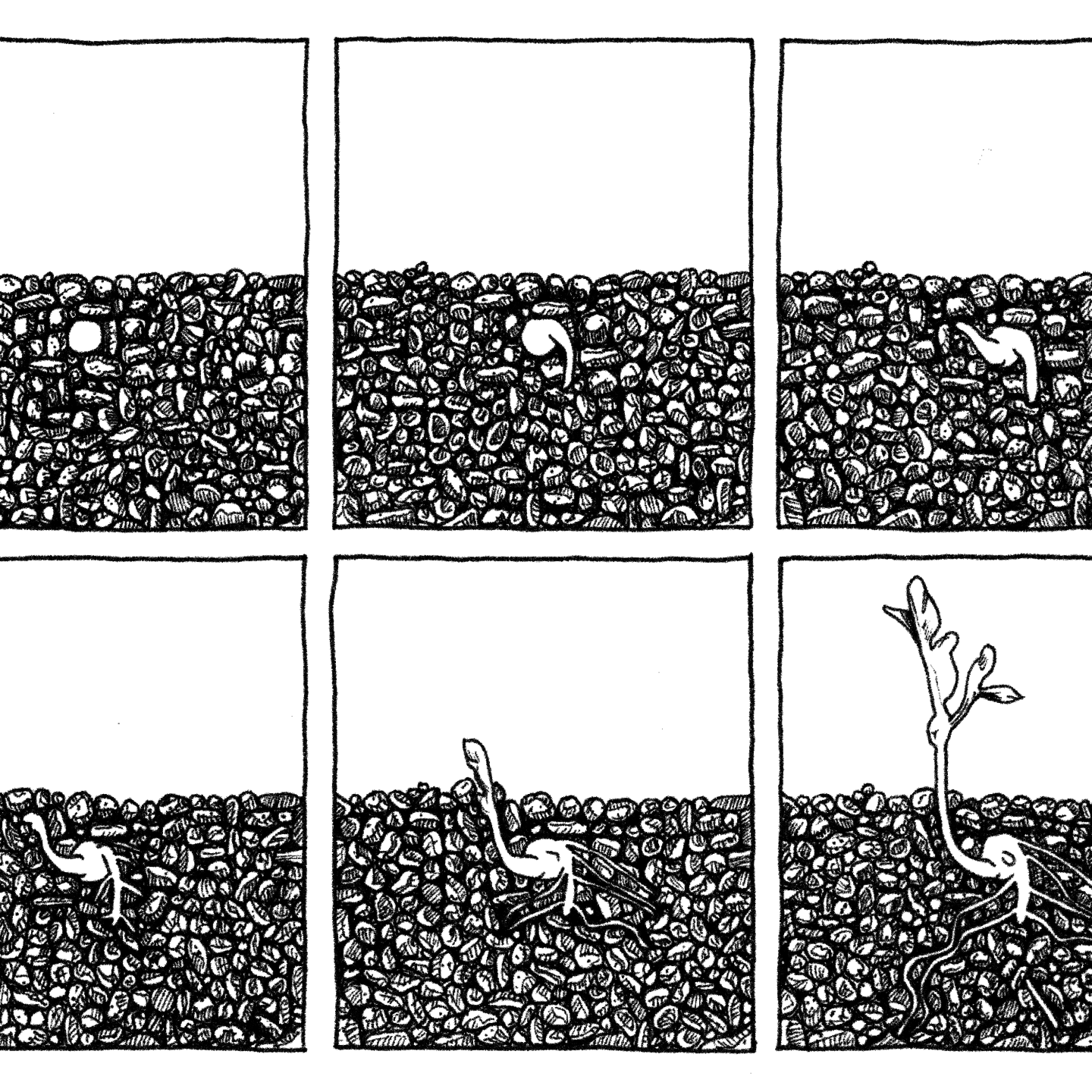
In What a Plant Knows, Daniel Chamowitz reports what plant biologists apparently have known for a long time: although plants generally stay in one place (they’re sessile), they actively negotiate their environments. Not just their cells, like all living cells, constantly do things, but whole plants and their parts—their roots, shoots, and leaves—respond to their environment.
For instance, plants use sunlight to convert water and carbon dioxide into energy. But plants don’t just get or fail to get sunlight. Most seek it. They grow towards it. If light illuminates one side of an oat seedling for long enough, that seedling will bend toward the light. If the light moves to the other side of the seedling, the seedling will in time bend in that direction. This is phototropism. A tropism is an oriented response to a stimulus. In phototropism, the plant does not merely respond to light—say, by getting warmer or starting photosynthesis—it responds to the direction of the light, either towards it (positive phototropism) or away from it (negative phototropism).
I have been talking of plants, as if they are all the same, but they are impressively diverse. There are mosses, liverworts, sequoias, and daffodils.
![]()
![]()
Contemporary botanists distinguish between vascular and non-vascular plants. Vascular plants have xylem, tissue dedicated to transporting water within the plant. Non-vascular plants don’t have xylem. Non-vascular plants, such as mosses and liverworts evolved first, before vascular plants, such as sequoias and daffodils.
The vast majority of plants are vascular. Their tissues are typically arranged concentrically, one sort of tissue nested within another, roughly like a series of cylinders. Cells of different tissues are different; some have stronger walls, providing support; others have special coatings, providing protection, or impeding movement of certain substances; others are for transporting substances, such as water. Most vascular plants have roots, shoots, and leaves. Roots anchor the plant, also supplying it with water and minerals absorbed from the soil. These are then transported to the rest of the plant through the xylem, tissues of dead cells that are roughly tube-like, connected end to end, extending upward through the shoot and outward to the branches. The xylem are commonly arrayed in a ring around the center (the pith) of the plant. Leaves are the primary site of photosynthesis, which generates food (glucose) for the plant. This food moves to the rest of the plant through the phloem, which is similar to the xylem, forming a ring of channels ordinarily positioned just outside the xylem. Together, the xylem and phloem count as the vascular system.
The majority of plants are vascular, bear seeds, and flower. A seed is a fertilized egg of a plant in a protective coating. Seeds allow plants to propagate across great distances, borne by wind or water or stuck to other organisms. Flowers are the reproductive parts of the plant, the sex organs. Typically, flowers have both “male” and “female” parts, ones that can produce a small gamete (pollen) and a large gamete (egg). Since flowering plants are the most common, it is appropriate for us to focus on them.
Now consider another tropism. Like most plants, a daffodil’s stem grows up out of the soil, while the roots grow down into it. Shoots go up; roots go down. If the whole plant grew down into the soil, it wouldn’t get enough or any sunlight. Roots need to be in the soil to tap water and nutrients, and to anchor the plant. Somehow these facts are integrated into these plant behaviors, which are instances of gravitropism, oriented responses to the direction of gravity. Roots exhibit positive gravitropism, growing with or towards the direction of gravity. Shoots exhibit negative gravitropism, growing against the direction of gravity.
![]()
Phototropism and gravitropism thus can tempt us to say that plants perceive.
Not so fast, you might say. True, plants respond to some aspects of their environment, but so too do rocks and puddles. Puddles evaporate when the temperature rises sufficiently, but they don’t perceive anything.
Maybe so, but plants are not merely responsive to their environments. Many of a plant’s responses have a goal, purpose, or point. Very roughly, phototropism has the goal of exposure to light. A puddle’s evaporating has no goal.
Fair point, you might admit, but human artifacts, such as a water wheel, have goals and they perceive nothing. A water wheel’s goal is to power a mill, but that doesn’t imply that in rotating in response to rushing water the wheel perceives that water or its rushing or anything at all.
Ah, here some of our initial questions sprout. Some non-organisms, such as water wheels, might indeed have goals, but they nevertheless differ importantly from plants. A water wheel does not aim to maintain itself. By contrast, a plant does aim to maintain itself. Furthermore, machines and plants have their goals in different ways—exogenously and endogenously, we might say. A water wheel’s goal is imposed on it by something other than itself. If the wheel goes awry, the miller addresses it. A plant’s goal is not imposed on it by something other than itself. If its shoots or roots go awry, the plant itself addresses it.
The urge to say plants perceive is rooted not simply in their responsiveness, or even their goal-directed responsiveness. Rather, some plant responses plausibly qualify as perceptions because they serve self-maintenance in a changing environment.
“Many of a plant’s responses have a goal, purpose, or point” no they don’t just as ducks don’t have webbed feet in order to swim, that these aspects/attributes have proved a good enough fit for their environs to allow for such traits to become passed along and even amplified does not give them a telos.
Thanks, dmf. This is certainly a topic of contention among philosophers. Some, such as Ruth Millikan, say that when a trait is “selected for” it has a function or purpose. You reject that line of thought. Others, such as Robert Cummins, say that evolutionary history aside, webbed feet and phototropism still have a function simply because they play a specific in a larger system or activity. Do you also reject that line of thought? Do you have a preferred story? Or are you doubtful more generally about things having telos-es?
hi Chauncey, not sure how technical/particular your use of “function” is here (will have to look up RC thanks) if it just means something akin to mechanical (writ large) function (as in a heart like a pump) within a system/organism than that would be hard to argue with (as part of a human description of such things) but yes I would say that the having of telos (teloi) is something I’m generally doubtful about (increasingly even in terms of humans but that’s another matter).
One might think that distally is a important feature of bona fide perceptual systems (or at least a ubiquitous feature of paradigmatic cases of perceptual systems). Genuine perceptual systems don’t merely respond to ambient energy, but have multiple sensory receptors that allow for some degree of spatial resolution which can be used to track distal objects. The fact that plants respond to ambient energy in a coordinated fashion doesn’t seem to imply that there are multiple sensory receptors that can detect either the direction of the energy (e.g., light) or provide any degree of spatial resolution.
Thanks, Ryan. Yes, distality (I believe that’s what you intended with “distally”) is an interesting feature of some paradigmatic perceptual systems, especially vision. One initial worry is that taste, smell, and touch which seem to be perceptual faculties, don’t exhibit it. (Perhaps here you would say that taste, smell, and touch are genuinely perceptual but only sensory.) Tyler Burge’s *Origins of Objectivity* is a thoughtful meditation on the significance of so-called perceptual constancy. For him, I don’t think it matters very much whether there are multiple sensory receptors or only few.
Separately, I admit that plant responses don’t seem to involve “spatial resolution”. Still, plants do more than respond to energy in an organized fashion. The organization of the responses takes a very special form.
Why would you say that taste, smell, and touch (in the paradigm cases) don’t exhibit distality?
Thanks, Ryan. We should probably get clearer about distality. What is it for some sensory ability to ‘exhibit distality’?
Although we humans are able with the aid of other sensory abilities and conceptual thinking to distinguish between, the source of the (proximal) stimulation and the source itself, the sense of taste and touch don’t seem to do this all on their own. Smell might do it more so. Put another way, they don’t distinguish between sources of stimulation as nearer or as farther. These senses are responsive to the proximity of the stimulus, but the senses themselves don’t seem to distinguish to distal stimulus from proximal effect (on the organism). Vision and audition seem to be interestingly different in that respect.
(In case your curious, Burge addresses this topic in relation to “objectification” and constancy on pp.415-416 of *Origins of Objectivity*. I’m sympathetic with what he says there.)